
Salt constructs in paleo-lake basins as high-priority astrobiology targets.
 1,
1,- 1Johns Hopkins Applied Physics Laboratory, United States of America (michael.phillips@jhuapl.edu)
- 2SETI Institute Carl Sagan Center, Mountain View, CA, United States of America
- 3NASA Ames, Space Science Division, Moffett Field, CA, United States of America
- 4Department of Geosciences, University of Montana, Missoula, MT, United States of America
- 5Department of Earth and Planetary Sciences, University of Tennessee, Knoxville, TN, United States of America
Introduction
In environments where it is difficult for life to function, microbial organisms tend to inhabit pockets of locally favorable climatic conditions. Micro-climates conducive to the persistence of life in an otherwise inhospitable environment – “refugia” – are spatially restricted and can be < centimeters in extent [1], [2]. Refugia may have been (and perhaps still are) perennially prevalent on Mars where conditions were likely never globally favorable to life for sustained periods of time [3]. The tendency for refugia to be small means that it may be difficult to locate features that could have served (or perhaps still do serve) as refugia for microorganisms on Mars. The spatial distribution of refugia in extreme environments across larger geographic extents is often non-random and may depend on many factors, biotic and abiotic [4]–[6]. Understanding patterns that refugia follow across larger geographic contexts as well as particular geologic phenomena (e.g., volcanic vents, dikes, stress fields) that are commonly associated with refugia may provide a way to infer regions of astrobiological interest, even if the specific, small, habitable patches (refugia) are below the resolving power of orbital instruments [6]. Here, we explore a case study of two terrestrial habitats in salt-encrusted paleo-lake basins (salars) in the Atacama and Altiplano of Chile to assess their characteristics and what factors are common between them. The Neogene salars of the Atacama and Altiplano are perhaps the best analogs on Earth for the Noachian/Hesperian salt-encrusted paleo-lakes of Mars [4], [7]–[12].
Evaporite habitats at Salar Grande and Salar de Pajonales
Salar Grande hosts decimeter scale nodules made of halite that serve as refugia for endolithic microbes [9]. [1] proposed a model to describe the evolution of nodules in halite-encrusted salars at the edges of polygonal features. To briefly summarize their model, halite nodules initiate at polygon edges in a salar with active ground water. Growth continues after ground water activity ceases as winds drive a moisture gradient, along which brines travel, toward the apex of the relatively higher relief nodules where more halite is deposited (Fig 1). The action of brines in halite nodules generates porosity at multiple spatial scales (nanometers to millimeters), contributing positively to their habitability [1].
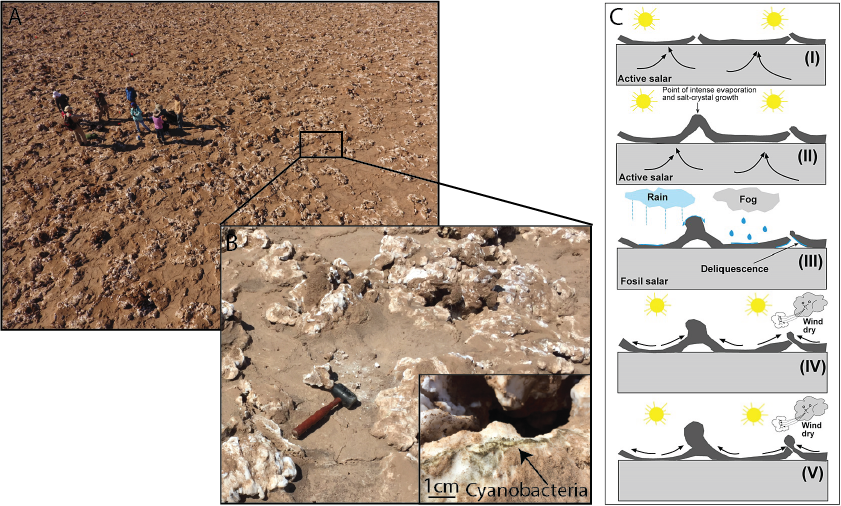
Fig. 1 Halite nodules at Salar Grande. A) Drone-view of nodules and nodule clusters. Humans for scale. B) Close up of halite nodule showing endolithic community. C) Halite nodule evolution from [1].
Like Salar Grande, Salar de Pajonales hosts endoliths in refugia habitats. In a gypsum-covered region of Salar de Pajonales, alabaster (a high-porosity polymorph of gypsum) is the most reliable indicator for the presence of life [4]. Alabaster refugia are most commonly found associated with decimeter- to meter-tall ridges and domes [6], [10]. The domes and ridges form via water-related processes: hydration/dehydration cycles, volume changes associated with mineral precipitation from brines, and/or efflorescence deliquescence [10]. The formation of alabaster is likely predicated on the action of the near-surface water that drives the formation of ridges and domes (Fig. 2), though microbial activity may play a role as well [4]. Therefore, at Salar de Pajonales water activity generates positive topographic salt constructs and physiochemical changes to gypsum (formation of high-porosity alabaster) that foster an environment favorable to life.

Fig. 2 Models of ridge and dome formation at SdP. A) Drone-perspective view of ridges at SdP. B) Model for ridge formation from [10] involving volume change at the phreatic-vadose zone interface. C) Image of domes at SdP in different stages of development. D) Possible model of gyspsum dome formation from [16].
Discussion
Across two salt-encrusted environments, one in the Atacama and the other in the Altiplano, with distinct evaporite mineralogy (halite versus gypsum), the activity of water resulted in decimeter- to meter-tall topographic constructs with nanometer- to millimeter-scale porosity conducive to the persistence of endoliths. We hypothesize that decimeter- to meter-tall topographic constructs (as opposed to erosional remnants or boulders) may be general indicators for relatively enhanced habitability in salt-encrusted paleo-lake basins because they require water to form. Although refugia – such as the precise location of endoliths in halite nodules or alabaster in gypsum domes and ridges – may not be observable from orbit, decimeter- to meter-scale salt constructs may be possible to identify with HiRISE or future orbital imagers with higher resolving power [6]. Chloride basins should be the targets of high-resolution imaging campaigns and efforts should be made to distinguish salt constructs from erosional remnants, boulders, and other relative topographic highs with which they could be confused. Salt constructs may be one of the few features, other than (fossil) hydrothermal vents, that have a high potential to both host and preserve microbial organisms, and that are specific targets, possibly identifiable from orbit, to which a rover could be driven. These characteristics make them attractive targets for future missions to Mars.
Reference:
[1] O. Artieda et al., 2015, doi: 10.1002/esp.3771.
[2] L. Hays, “NASA Astrobiology Strategy.” 2015.
[3] R. Wordsworth et al., 2021, doi: 10.1038/s41561-021-00701-8.
[4] K. Warren-Rhodes et al., Nature Astronomy. in review.
[5] M. S. Phillips et al., Astrobiology, in review.
[6] K. A. Warren-Rhodes et al., 2019. doi: 10.3389/fmicb.2019.00069.
[7] M. M. Osterloo, et al., 2010, doi: 10.1029/2010JE003613.
[8] T. D. Glotch, et al., 2016, doi: 10.1002/2015JE004921.
[9] A. F. Davila et al., 2008, doi: 10.1029/2007JG000561.
[10] N. W. Hinman et al., 2022, https://www.frontiersin.org/article/10.3389/fspas.2021.797591
[11] N. A. Cabrol et al., 2007, doi: 10.1029/2006JG000298.
[12] E. K. Leask and B. L. Ehlmann, 2022, doi: 10.1029/2021AV000534.
[13] A. Szynkiewicz, et al., JGR, vol. 115, 2010.
How to cite: Phillips, M., Warren-Rhodes, K., Hinman, N., Moersch, J., Hofmann, M., McInenly, M., Davila, A., and Cabrol, N.: Salt constructs in paleo-lake basins as high-priority astrobiology targets., Europlanet Science Congress 2022, Granada, Spain, 18–23 Sep 2022, EPSC2022-1188, https://doi.org/10.5194/epsc2022-1188, 2022.